This study was designed to assess sex-related differences in the
selection of an appropriate strategy when facing novelty.
A simple visuo-spatial task was used to
investigate exploratory behavior as a specific response to novelty.
The exploration task was followed by a
visual discrimination task, and the responses were analyzed using signal
detection theory.
During exploration women selected a local searching strategy in which
the metric distance between what is already known and what is unknown
was reduced, whereas men adopted a global strategy based on an
approximately uniform distribution of choices. Women's
exploratory behavior gives rise to a notion of a secure base warranting
a sense of safety while men's behavior does not appear
to be influenced by risk. This sex-related difference was interpreted
as a difference in beliefs concerning the likelihood of uncertain
events influencing risk evaluation.
Keywords: exploration, spontaneous strategies, sex differences,
decision-making.
Males and females seem to differ in spatial abilities and styles
(e.g., Jahoda, 1980; Mann, et al., 1990). Generally, studies
involving navigational problems showed that female cognitive style
relies more on detailed information, while male style relies more on
global information (e.g., Meyer-Levy, 1989; Moffat et al., 1998;
Silverman et al., 2000). Evolutionary mechanisms could potentially
account for sex differences in spatial behavior. For example, these
behavioral differences may be due to mating patterns that induced a
selection of large-range navigation in males (Gaulin & Fitzgerald,
1986, 1989). Mating patterns or mating strategies are linked to the
dynamics of reproduction and sexual selection (Darwin, 1871), and
sexual selection is restricted to characteristics that influence mate
choice and competition for mates. Typically, males have to compete
through extensive ranging for access to mates while females have to
choose mating partners according to reproductive success (for reviews:
Andersson, 1994; Trivers, 1972; Geary, 2000).
Another proposition, but exclusively directed at humans, suggested
that the division of labor (game hunting and plant gathering) would
have put greater selection pressure on females' spatial memory because
females sustained gathering duties (Silverman & Eals, 1992).
However, as argued by Ecuyer-Dab and Robert (2004a),
"the selection of male characteristics depends on
females' choice for mates. In females, however, spatial
cognition would have been primarily shaped by the natural selection of
a strong concern for survival (both of self and of offspring). This
concern would have compelled them to favor low-risk strategies, like
concentrating on proximal spatial cues, when coping with space-related
problems. Such focusing would have enabled secure navigation based on
detailed landmark encoding, as well as, in certain species, regular
feeding based on remembering the exact locations of potential
resources" (p. 222). Thus, the hypothesis of labor
division would be a by-product of sexual selection and not the cause
of sex differences in spatial behavior.
Taken together, the literature seems to indicate that the key to
understanding the evolution of behavioral sex differences relies on the
relative costs and benefits of producing offspring (see Trivers, 1972,
but also Geary, 2000). In that context spatial skills play a crucial
role since they increase reproductive success and the accessibility to
food resource but, at the same time, multiply the risks of getting
lost, being killed or consumed by other animals (predation). Hence, the
survival of mobile species depends on their ability to balance costs
and benefits induced by locomotion and this balancing should differ
according to sex.
Experimental investigations of sex differences in spatial abilities
yield apparently disparate results (e.g., Jones et al., 2003;
Ecuyer-Dab & Robert, 2004a). This might be partly due to the
complexity of contemporary experimental designs, but also to a lack of
investigations concerning decision-making processes involved in the
selection of strategies.
The current study investigates sex differences in basic behaviors
like exploration, detection and discrimination involving the selection
of strategies when coping with uncertainty. Following the above
quotation from Ecuyer-Dab and Robert's, the hypothesis is that women,
compared to men, should favor low-risk strategies when
coping with space-related problems. To test this, I used a
simple spontaneous two-dimensional exploratory task. This choice relies
on the fact that exploration is a natural behavior and that it is
fundamental in acquiring spatial knowledge. It seems to be based on
driving factors such as curiosity, comfort or mastery over
one's environment. Moreover, it is commonly defined as
serving to reduce uncertainty and thus allow coping with fear (for a
review about exploration, see Hughes, 1997).
Exploration is mainly characterized by a succession of progressions and
stops (Drai, Benjamini, & Golani, 2000), and the selection of exploration
could rely on its capacity to act as a regulator of uncertainty.
Indeed, progressions are based on decisions taken during stops, and
stops correspond to choice points allowing decisions. Voss (1983)
refers to the exploration process as the generation and testing of
hypotheses concerning the object's meaning and
potential use.
In order to assess risk-taking, a classical visual discrimination task
based on the stimuli observed during the exploration task was used. The
results were analyzed with signal detection theory.
The participants were 160 volunteers (80 males and 80 females) recruited
from the campus of the University of Lausanne (Switzerland). The mean
age of the males and females was 22.9 (SD 1.4) and 23.5 (SD 1.2) years,
respectively.
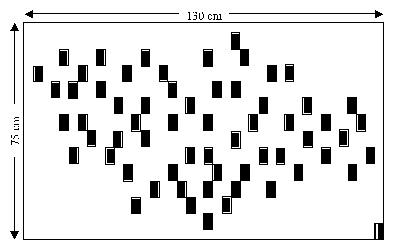
Figure 1: Two-dimensional pattern used for Experiment 1, the card placed
at the lower right corner corresponds to the goal.
A set of one-hundred cards differentiated by homogenous male and female
characters was used.
For Experiment 1, a two-dimensional pattern (Figure 1) was created in
placing 62 cards (3 x 5 cm) on a large board (130 cm x 75 cm). Two men
and two women designed the shape of the pattern to avoid any sex bias.
A card was randomly chosen and duplicated to serve as goal. The
remaining set of 38 cards was used for Experiment 2.
The board lay on the floor of the experimental room with the 62 cards
turned the wrong side up. The duplicate of the goal turned right side
up was put on the lower right corner of the board (Figure 1). Each
participant was greeted and placed in front of the board and was asked
to find the hidden goal by turning up as few pictures as possible and
leaving them exposed. After having found the goal, participants were
asked to turn up the remaining pictures in order to look at all
characters. Double blind shuffling of cards made different strategies
(e.g., systematic or random) equivalent with regard to the probability
to get the goal in one draw.
A sample of 32 new pictures of characters was added to the original
sample of 62 pictures used during the exploration phase and the
hundred pictures (62 already shown in Experiment 1 and 38 new) were
randomly presented one by one to the participants. The participant's
task was to discriminate between the pictures already shown (signal)
and the one never shown during exploration. Subjects' responses were
coded as followed: 1/ hit (present, already shown, correct response),
2/ false alarm (present, never shown, incorrect response), 3/ correct
rejection (absent, never shown, correct response), 4/ miss (absent,
already shown, incorrect response).
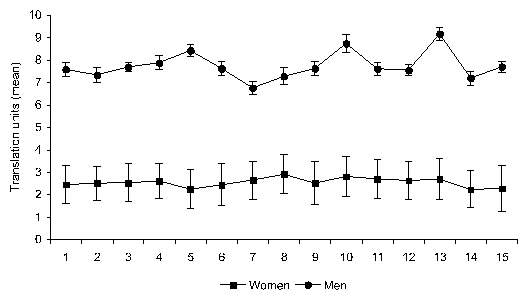
Figure 2: Mean ( sem) axial translations during the first
fifteen draws (mean women: 2.56 0.09; mean men:
7.73 0.26)
Exploration was assessed through axial translations units between
explored locations during the first fifteen draws. As shown in Figure
2, men adopted a global strategy in which pictures were randomly
chosen from an approximately uniform distribution over spatial
locations. In contrast, women used a local searching strategy in which
the probability of selecting a card was a function of its distance
from a cluster of visited locations. Difference in searching strategy
was confirmed by a one-way ANOVA on mean units translation
(F(1,158)=372.95; p=.001) indicating that the mean unit translation
was larger in men than in women (Fisher's PLSD men
women, p=.001).
This difference in strategy is quite intriguing and might be related to
risk perception. Risk can be seen here as a
chance of something negative happening. In the
present task, finding the goal by turning up as few picture as possible
has been put at stake. Thus, the probability of not being successful on
the first trial was high and risk evaluation should be high too. Early
psychologists (Angell, 1907; Fernberger, 1914; Boring, 1920) argued
that uncertainty biases responses because it alters confidence, moods
and temperament (insecurity or boldness for example). Analogically,
exploration of unfamiliar environmental stimuli by a free moving animal
has been described as competing states of both fear and curiosity
(Montgomery, 1955). In this case, uncertainty and fear, with respect to
the external environment, appears to be reduced when the present
environment is compared with previously experienced stimuli (Russell,
1983). Another component made apparent by the study of risk
assessment behavior (new object reaction) in animals was how
significant it is for these animals to keep continual contact with
familiar markers (Blanchard et al., 1970; Blanchard & Blanchard,
1989; Misslin & Ropartz, 1981). There is a parallel between these
animal behaviors and the searching strategy adopted by women. The known
part of the board could be used as a secure base and minimizing the
distance between this base and what is unknown could permit to feel
safer while exploring the environment. Indeed it allows balanced
exploration and seeking the proximity of familiar markers. Under this
experimental condition, the behavior of women might be interpreted as a
risk-reducing strategy allowing coping with uncertainty. To support
this hypothesis, it must be added that usually, but not in this task,
nearby environments are likely to be more similar in the probability of
what they contain. Similarly, distant environments are less known,
more uncertain in the sense of having missing information.
However, an alternative hypothesis must be considered. It has been
postulated that spatial skills depend on the amount of spatial
information processed according to usual home range (Gaulin and
Fitzgerald, 1986, 1989). Thus, women's search pattern could be in
relation with this range size navigation mode hypothesis
(Ecuyer-Dab & Robert, 2004b) since they have a tendency to focus on a
smaller-scale space than men. Accordingly, differences observed in
searching strategies could depend on differences in the selection of
environmental features to guide behavior. Indeed, it has been shown
that men rely primarily on cardinal references and geometrical
environmental features while women rely more on topological cues such
as landmarks and their configurational relations (e.g., Montello et
al., 1999; James & Kimura, 1997; Choi & Silverman, 1996).
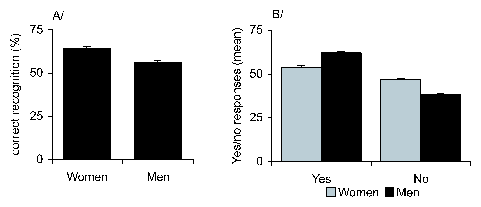
Figure 3: A. Mean ( sem) of "yes" and "no"
responses in men and women during the discrimination task. B. Mean
( sem) percentage of correct recognition by men and women
during discrimination.
Taken together, these hypotheses suggest that at least two main
cognitive processes control exploration and spatial behavior: 1/
Information processing that extracts, selects and encodes relevant
information provided by internal and external worlds; 2/ decision or
choice processes allowing selecting a strategy. These two operations
are altered, respectively, by attentional mechanisms that change
discrimination capacities, and by beliefs concerning the likelihood of
uncertain events. Information processing is tuned by the attentional
level that acts like a filter on perception, while decision-making
processes are weighed by subjective probability
of risk. Consequently, strategies selected during
exploration should be mainly controlled by these two mechanisms.
In order to investigate sex differences in these two cognitive
processes, a classical visual discrimination task based on the stimuli
observed during the exploration task was used.
Analysis of responses showed that men said more "yes" and less
"no" responses than women (multivariate analysis sex x yes:
F(1,158)=52.16; p=.001, sex x no: F(1,158)=53.3; p=.001), but the
percentage of correct recognitions, assessed through a classical
coding of answers as correct and incorrect responses, was
significantly higher in women than in men (F(1,158)=35.77; p=.001;
Fisher's PLSD women men, p=.001) as shown in Figure 3.
The analysis of the percentage of correct recognition
does not allow dissociating between perception and judgment. Signal
detection theory (SDT) provides a tool for quantifying perceptual
decisions in the presence of uncertainty (Green & Swets, 1966). This
theory treats detection of a stimulus as a decision-making process
determined by the nature of the stimulus and cognitive factors.
The model generally used in SDT assumes that the theoretical
distributions of signal and noise are normal and have equal variance.
The probability of correct and incorrect signal detection can be
calculated from the ratio of the subject's acceptation
and rejection responses (Table 1). This probability is then used to
determine the probit transformations aimed at estimating
d' and c (Green & Swets, 1966).
Table 1: Calculation of the probabilities of correct and
incorrect detection of a signal (S= signal; N= noise; A= absent; P=
present).
Responses:
Signal:
present
absent
Present
S
P/S
Hit rate
A/S
Miss rate
Absent
N
P/N
False alarm rate
A/N
Correct rejection rate
The d' index correspond to the distance
between means of the distributions. It measures the strength of the
stimulus relative to the sensitivity of the subject. The c index
corresponds to the likelihood ratio. It reflects the strategy of the
subject and can be changed with level of confidence. A low c matches
with a rash temperament while a high c matches with a conservative
temperament.
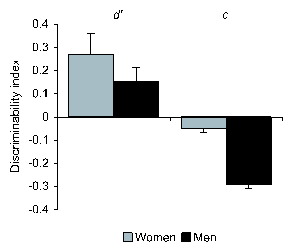
Figure 4: d' and c index values (mean
sem) measuring sensitivity and temperament in women and men.
The SDT analysis showed that the sensitivity
d' did not significantly differ between men and women
(F(1,158)=1.33; ns), but the subjective criterion c was significantly
different (F(1,158)=75.74; p=.001). According to the theory, this last
result indicates that women's decisions were, on
average, more circumspect and were based on conservatism while
men's decisions were more risky, in the sense of risking false
positives (Figure 4).
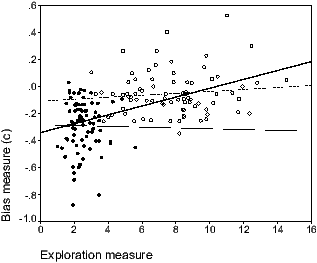
Finally, correlations as a whole, and
within women and men between the exploration measure and the bias
measure (c) were calculated. As shown in Figure 5, the overall analysis
revealed a significant correlation between these two measures
(r(160)=.498; p=0.01). However, separate analyses conducted
within each sex were not significant (women: r(80)=-.012; ns;
men: r(80)=.112; ns).
Figure 5: Relation between the exploration measure and the bias measure
in women and men. Exploration measure corresponds to the mean
translation unit observed during the exploration task, whereas the bias
measure corresponds to the c index of the discrimination task.
This study has first shown that the searching strategies used by women
and men to solve an exploratory task that may be seek as involving trading off of risk and
reward differed according to sex. Women adopted a local searching
strategy in which the metric distance between what is already known
and what is unknown was reduced. Men adopted a global strategy based
on an approximately uniform distribution of choices. These findings
appear to be compatible with a female frame of mind expressing careful
consideration of all circumstances and possible consequences before
making a decision.
The results of the discrimination task through a signal detection
analysis showed that women and men sensitivity towards the stimuli
(d' index) was equivalent while the assessment of confidence
in judgment under uncertainty (c index) differed. Indeed, men showed a
subjective criterion indicating a more risky temperament than women.
The absence of differences in sensitivity contrasted with what is
generally expected from results showing that women outperform men when
the task requires object identification (Silverman & Eals, 1992;
James & Kimura, 1997) or item memory (Galea & Kimura, 1993,
Maccoby & Jacklin, 1974, Stumpf & Jackson, 1994). If correlation
as a whole supported the hypothesis that strategies selected during
exploration rely on temperament, it was not the case for correlations
within each group.
It could be argued that these measures were not based on the same
cognitive mechanism. Indeed, subjects were facing with the unknown in
the exploration task while having to decide if they had already seen or
not a picture in the discrimination task. Thus, these two tasks might
not be equivalent according to uncertainty but also according to
one's confidence in skills. For example, LaGrone (1969)
reported that women more often felt disoriented with regard to their
"sense of orientation" than men did.
Moreover, women expressed a feeling of "worry about
becoming lost" that was negatively associated with
confidence in the "sense of orientation"
(Kozlowski & Bryant, 1977). It has also been shown that the
positive representation of an activity in term of payoffs and costs
influenced women's favorable judgments concerning
consequences (Harris et al., 2006). Interestingly, variations in the
relation between exploratory behavior and risk-taking have been also
observed in birds (Parus major). In this species, correlation between
exploratory behavior and risk-taking behavior depended on the presence
and foraging behavior of a mate. Moreover, the presence of a mate
resulted in a complex differential effect according to the fact that
they were males or females and slow or fast explorers. Slow explorers
of both sexes were more audacious in the presence of a mate whereas in
fast explorers, females were less audacious in the presence of a mate
(van Oers et al., 2005).
The results of this experiment, together with those of an abundant
literature, cannot lead to a clear understanding of why and how the
mentioned abilities are associated with beliefs concerning the
likelihood of information and degrees of confidence consistently with
sex differences. One possibility to be examined is that
sex-steroid hormones could modulate cognitive mechanisms involved in
risk evaluation.
Finally, it is perhaps of some interest to note that the present work
was drawn from Ittelson (1960) suggestion that "the
problem of exploratory behavior... [is] central to the study of
environmental perception."
It differs from previous works since it offers a new perspective to
assess sex-related differences in spatial behavior through its relation
to perception. Perception and sensation should be regarded as
particular forms of extended communicative actions and not as
alternative end points for the use of environmental information (Green,
2001). All information about the world comes from our sensory systems,
and the cognitive process begins with the ability to make some meaning
out of this stream of sensory stimuli.
References
Andersson, M. (1994). Sexual selection. Princeton, NJ,
Princeton University Press.
Angell, F. (1907). On judgment of "like" in
discrimination experiments. American Journal of Psychology,18, 253.
Blanchard, R. J., Shelton, V. T. and Blanchard, D. C. (1970). Historical
effects of stimulus exposure: Readiness to eat and object exploration.
Learning and Motivation, 1, 432-444.
Blanchard, R. J. and Blanchard, D. C. (1989). Attack and defense in
rodents as ethoexperimental models for the study of emotion.
Progress in Neuro-Psychopharmacology & Biological Psychiatry,
13, 3-14.
Boring, E. G. (1920). The control of attitude in psychophysical
experiments. Psychological Review, 27, 440-452.
Choi, J., and Silverman, I. (1996). Sexual dimorphism in spatial
behaviors: applications to route learning. Evolution and
Cognition, 2, 165-171.
Darwin, C. (1871). The descent of man, and selection in relation
to sex. London: John Murray.
Drai, I., Benjamini, Y., Golani, I. (2000). Statistical discrimination
of natural modes of motion in rat exploratory behavior. Journal
of Neuroscience Methods, 96, 119-131.
Ecuyer-Dab, I. and Robert, M. (2004a). Have sex differences in spatial
ability evolved from male competition for mating and female concern for
survival? Cognition, 91, 221-257.
Ecuyer-Dab, I., and Robert, M. (2004b). Spatial ability and home range
size: examining the relationship in Western men and women.
Journal of Comparative Psychology,118(2), 217-231.
Fernberger, S. W. (1914). The effects of attitude of the subject upon the
measure of sensitivity. American Journal of Psychology,25, 538-543.
Galea, L. A. M. and Kimura, D. (1993). Sex differences in route-learning.
Personality and Individual Differences, 14, 53-65.
Gaulin, S. J. C. and Fitzgerald, R. W. (1989). Sexual selection for
spatial-learning ability. Animal Behaviour, 37, 322-331.
Gaulin, S. J. C., and Fitzgerald, R. W. (1986). Sex differences in spatial
ability: An evolutionary hypothesis and test. The American
Naturalist, 127(1), 74-88.
Geary, D. C. (2000). Evolution and proximate expression of human paternal
investment. Psychological Bulletin, 126, 55-77.
Green, P. R. (2001). The Relation Between Perception and Action: What
Should Neuroscience Learn From Psychology? Ecological
Psychology, 13, 117-122.
Green, D. M. and Swets, J. A. (1966). Signal detection theory and
psychophysics. New York, Wiley.
Harris, C. R., Jenkins, M. and Glaser, D. (2006). Gender differences in
risk assessment: why do women take fewer risks than men?
Judgement and Decision making, 1, 48-63.
Hughes, R. N. (1997). Intrinsic exploration in animals: motives and
measurement. Behavioural Processes, 41, 213-226.
Ittelson, W. H. (1960). Visual space perception. Springer, New
York, p13.
Jahoda, G. (1980). Sex and ethnic differences on a spatial perceptual
task: Some hypotheses tested. British Journal of Psychology,
71, 425-431.
James, T. W. and Kimura, D. (1997). Sex differences in remembering the
location of objects in an array: location-shifts versus
location-exchanges. Evolution and Human Behavior, 18, 155-163.
Jones, C. M., Braithwaite, V. A. and Healy, S. D. (2003). The evolution of
sex differences in spatial ability. Behavioral Neuroscience,
3, 403-411.
Kozlowski, L. T. and Bryant, K. J. (1977). Sense of direction, spatial
orientation, and cognitive maps. Journal of Experimental
Psychology (Human Perception), 3, 590-598.
LaGrone, G. W. (1969). Sex and personality differences in relation to
feeling for direction. Journal of General Psychology,81, 23-33.
Maccoby, E. E. and Jacklin, C. N. (1974). The psychology of sex
differences. Stanford, CA. Standford University Press.
Mann, V. A., Sasanuma, S., Sakuma, N. and Masaki, S. (1990). Sex
differences in cognitive abilities: a crosscultural perspective.
Neuropsychologia, 28, 1063-1077.
Meyers-Levy, J. (1989). Gender differences in information
processing: a selectivity hypothesis. In Cafferata, P. C. and Tybout,
A. M. (Eds). Cognitive and affective responses to advertising. Lexington
Books, 219-260.
Misslin, R. and Ropartz, P. (1981). Effects of metamphetamine on
novelty-seeking behavior by mice. Psychopharmacology,
75,39-43.
Moffat, S. D., Hampson, E., and Hatzipantelis, M. (1998). Navigation in a
"virtual" maze: sex differences
and correlations with psychometric measures of spatial ability in
humans. Evolution and Human Behavior, 19, 73-87.
Montgomery, K. C. (1955). The relation between fear induced by novel
stimulation and exploratory behavior. Journal of Comparative
Physiology and Psychology, 48, 254-260.
Montello, D. R., Lovelace, K. L., Golledge, R. G., and Self, C. M. (1999).
Sex-related differences and similarities in geographic and
environmental spatial abilities. Annals of the Association of
American Geographers, 89, 515-534.
van Oers, K., Klunder, M., Drent, P. J. (2005). Context dependence of
personalities: risk-taking behavior in a social and a nonsocial
situation. Behavioral Ecology, 10.1093(ari054), 716-723.
Russell, J. A. (1983). Pancultural aspects of the human conceptual
organization of emotions. Journal of Personality and Social
Psychology, 45, 1281-1288.
Silverman, I., Choi, J., Mackewn, A., Fisher, M., Moro, J., and
Olshansky, E. (2000). Evolved mechanisms underlying wayfinding: further
studies on the hunter-gatherer theory of spatial sex differences.
Evolution and Human Behavior, 21, 201-213.
Silverman, I. and Eals, M. (1992). Sex differences in spatial abilities:
Evolutionary theory and data. In Barkow, J. H., Cosmides, L. and Tooby,
J. (Eds), The adapted mind: Evolutionary psychology &
generation of culture. NYC: Oxford Universities Press, 533-549.
Stumpf, H. and Jackson, D. N. (1994). Gender-related differences in
cognitive abilities: evidence from a medical school admission testing
program. Personality and Individual Differences, 17, 335-344.
Trivers, R. L. (1972). Paternal investment and sexual selection. In B.
Campbell (Ed.), Sexual selection and the descent of man
1871-1971 (pp. 136-179). Chicago, Aldine.
Voss, H.-G. (1983). Neugier und Exploration. In Euler, H. A., Mandl, H.
(Hrsg.), Emotionspsychologie: Ein Handbuch in
Schlüsselbegriffen, 220-226. Urban & Schwarzenberg, München.
Footnotes:
1Thanks are due to Jason Borioli for careful
reading. Address: Institute of Psychology, SSP
Faculty, Anthropole, CH-1015 Lausanne, Switzerland.
E-mail address: Catherine.Brandner@unil.ch.
File translated from
TEX
by
TTH,
version 3.78. On 24 Oct 2007, 10:37.